Let’s do a coronavirus post today, since there are some really good data out on the structure and function of the by-now-universal Omicron variant. As we have all been hearing, this one has a great number of mutations in the Spike protein region – via Eric Topol on Twitter, here’s a visualization from Nextstrain of where Omicron sits in phylogenetic space compared to previous variants, annotated by Emma Hodcroft at Bern. You can see that it’s an odd one, as advertised, pretty much off by itself and with its most recent common ancestor apparently coming from back in 2020 somewhere.
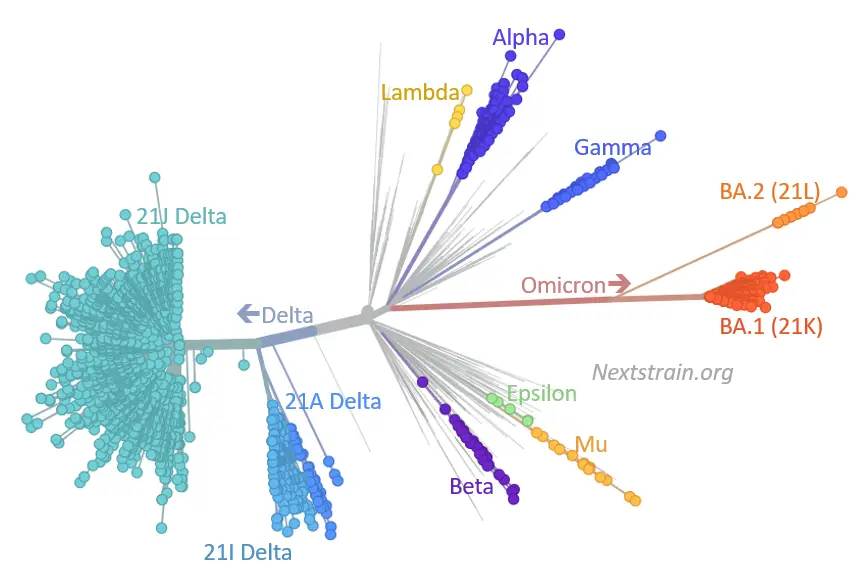
Nature has a good new article on the various theories for how this happened. That long arm poking out into Omicron territory on that phylogenetic tree tells you that we really didn’t pick up on any of the (many) intermediate mutations along the way. It’s possible that we just missed them in the wild as they spread from person to person, but a lot of people don’t quite believe that. So it could have developed inside an immunocompromised patient whose body kept fighting it to a roughly even draw for months, allowing the virus to be under selection pressure the whole time without killing it off. Or it could have been a jump (back in 2020) into another species, with evolution there until it made the jump back into humans. My own guess is the immunocompromised patient, or maybe more than one. I think that the other explanation with multiple interspecies jumps is kind of a stretch, but we really don’t have enough information yet to say anything with lots of confidence. It would be very good to know the answer, though, because we might then start some different forms of surveillance to keep from being taken by surprise again.
So what do all these mutations do for the virus? The first two links above gives us a few of those answers. The first paper has some very nice cryo-EM structural data of the Omicron virus and its binding to the human ACE2 protein, the second has similar data obtained via X-ray crystallography. This lets us see what at least a few of those mutated residues are doing. At right is an illustration from the cryo-EM paper showing the mutations in the Spike protein itself, shown as red balls. You can see a bunch of them in the actual receptor-binding domain (RBD), and some of these (like N501Y, T478K, and K417N) we’ve seen in other lineages.
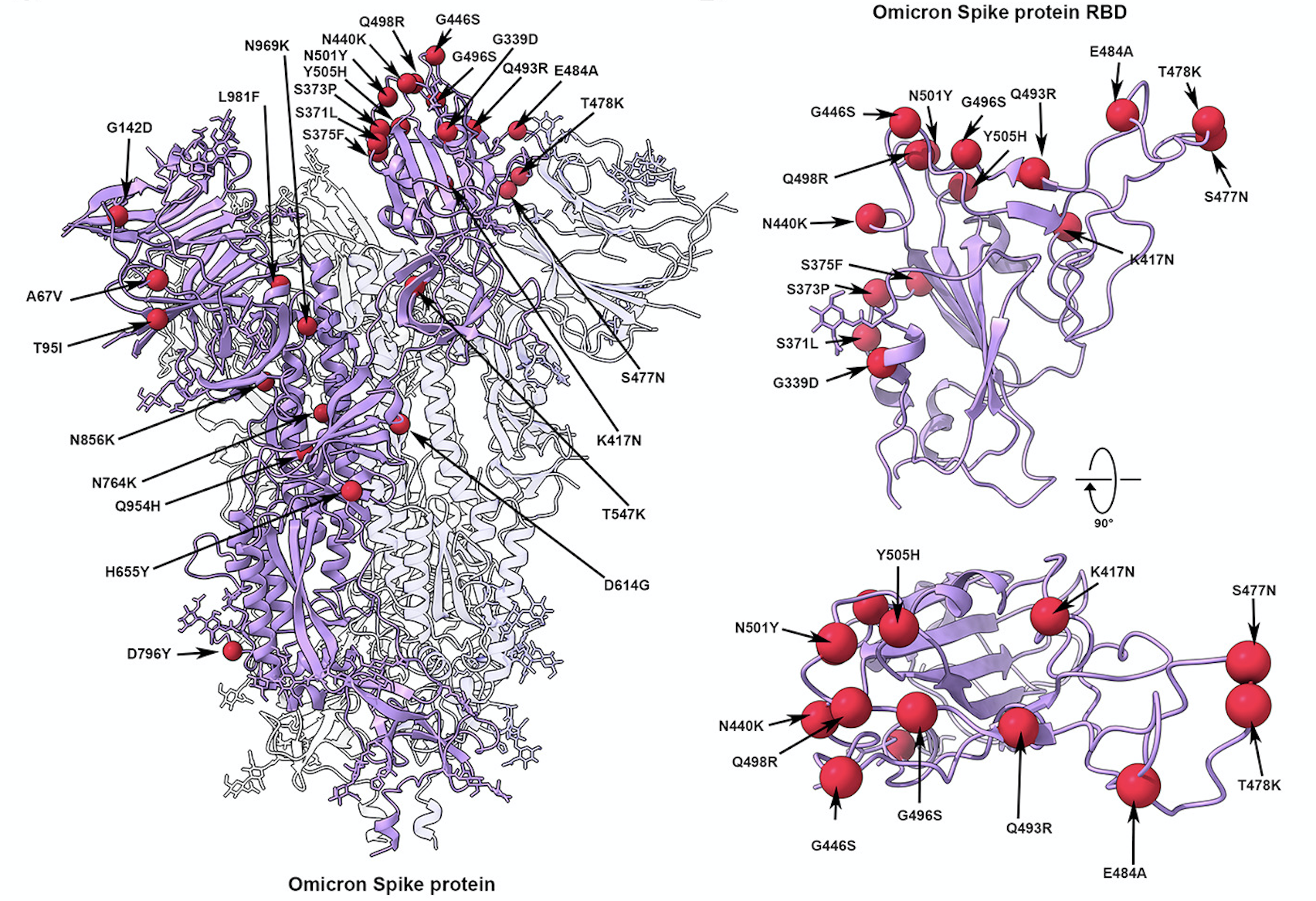
For those outside the protein world, that notation is how we usually write mutated residues. Each protein has a numbering system, starting from the end that has a free amino group on it (the N-terminal) and going to the end that has a free carboxylic acid on it, the C-terminal. Every amino acid in the protein is numbered in sequence, so you can see that those three residues just mentioned are four or five hundred amino acids down from the N-terminal. And the single letters are the amino acid code that’s used by molecular biologists and protein scientists around the world, as seen in this table. So when you say “K417N”, that means that the lysine (K) that’s normally at position 417 of the protein has been mutated to an asparagine (N). This information will make you a hit at parties, trust me, no matter how unappealing you may be otherwise.
The overall binding of Omicron’s spike is about the same as Delta, so that’s not what makes it special. In fact, a number of the mutations it has actually decrease its binding affinity, but those are made up for by others. The Q493R, G496S and Q498R residues seem to be doing that job, with those new arginines and the serine forming new salt bridges and hydrogen bonds. Meanwhile, that K417N mentioned above loses a nice salt-bridge interaction that was present in Delta.
So what are these mutations doing, if they’re not making the Spike bind its target better? You guessed it: evading the usual antibodies to a greater degree. That’s an argument for the chronic-infection-in-some-immunocompromised-patient theory, because this is what you’d expect to see after a long slugfest with human antibodies that could bind the surface of the virus but weren’t quite enough to finish the infection off. The authors of both papers linked above conclude that the virus has been walking an evolutionary tightrope between antibody evasion and retaining binding to ACE2, and unfortunately hit on a combination that allowed it to do both, and this fits in with earlier work on the Omicron genome as well. The X-ray paper also picked up on a new surface modification on N370, a glycan reside attached to that asparagine, which can also help with the immune evasion as it sits in the binding footprint common to many antibodies.
This helps to explain the number of “breakthrough” infections of Omicron in previously-vaccinated people, but as soon as you say that you have to follow up with the fact that the vaccines also have done a remarkable job of keeping people out of the hospital, not to mention the ICU or the morgue. In country after country where these data have been collected, the previously unvaccinated have had remarkably greater chances of ending up with a serious infection from Omicron. We have been fortunate that the immune response brought on by vaccination has performed that well against a variant like this one that has a really long list of mutations compared to the original vaccination version of the Spike. Remember, when you say “immune evasion” you’re not saying “total immune evasion” – I think many people have been confused by that point. Vaccination (and prior infection) raise a whole host of different antibodies, some of which are rendered less effective by Omicron’s mutation profile and some of which aren’t. People who have had the actual coronavirus can get up to even higher and more effective immunity by getting vaccinated on top of that.
And this also explains the way that Omicron has wiped out the effectiveness of some of the existing monoclonal antibodies (such as those from Lilly and Regeneron). This has recently become a political issue, with Florida’s governor Ron DeSantis angrily protesting the FDA’s statements about these antibodies becoming useless for treating Omicron patients. Various blowhards amplified this to the point of accusing the government of a conspiracy against Florida residents and others who were being treated with these antibodies, since many of these patients had also chosen to remain unvaccinated. The technical term that sums up this point of view is “horseshit”. As opposed to the variety of antibodies that you would have from vaccination or prior infection, mAb therapy gives you only one or two, administered in large quantities from an outside i.v. And if the Omicron mutations keep those exact antibodies from binding to the virus, then you are just completely out of luck. That’s what has happened – it’s not a case of “Well, they should keep these on the market because they might still help someone”. No, Omicron is now over 99% of all the coronavirus infections in the US, and these exact monoclonal antibodies are useless against Omicron. That ship has sailed.
Want a wider suite of antibodies than you can get from being dosed with a single monoclonal? Get vaccinated. If you’ve been infected, of course, you have a nice wide variety as well, but you got that while risking all the possible problems with the disease, ranging from sudden turns toward serious infection all the way to “Long Covid”. None of which you will get from the vaccine. The risk/benefit for most adults seems extremely clear, which is why watching thousands of people die every day in a way that the huge majority of them could have prevented is so damned frustrating.
